专家点评Nature | 突破!张毅团队揭秘染色体之外环状DNA的前世今生
转自:BioArt植物
点评 | 钟波、舒红兵(武汉大学)
责编丨迦溆
作为生命的遗传物质,DNA可分为线性和环状两种。真核生物基因组染色体DNA以线性方式存在,而细胞器(线粒体和叶绿体)、绝大多数细菌、部分病毒基因组DNA则以环状形式存在。染色体之外环状DNA(eccDNA,extrachromosomal circular DNA)指在真核生物中发现的,染色体外的、非线粒体、非叶绿体环状结构DNA。eccDNA的发现(1964)【1】甚至早于广泛熟知的线粒体DNA【2-4】和细菌质粒DNA【5,6】,两者分别于1966年和 1967年被显微镜观察证实为环状。
历经多半个世纪的研究,人们对eccDNA与基因组DNA的序列高度同源,已经达成共识,但因其尺寸差异巨大:从数百bp(碱基对)到数百kb(千碱基对)不等,研究人员据此提出了多种命名,microDNA主要指400 bp以内的环状DNA【7】,ecDNA (extrachromosomal DNA)【8】被用以描述在癌细胞中发现的、尺寸数百kb以上的染色体外DNA,其巨大尺寸,足以容纳全长基因和DNA复制起始位点,因此能自主复制和扩增,并与癌基因的扩增和癌症的发生有关。 而eccDNA正被用来指比ecDNA小的所有染色体外环状DNA,这些eccDNA几乎存在于所有的真核细胞系和组织器官中,与染色体DNA相比较,其丰度极低,含量易变。迄今为止,关于eccDNA三大基本问题仍然没有答案:1,eccDNA与基因组的序列同源性是否具备特异性,(从哪来)?2,eccDNA的产生机制是什么(怎么来)?3,eccDNA有无生物学功能(做什么)?
2021年10月20日,哈佛医学院、波士顿儿童医院张毅教授实验室以长文的形式在Nature杂志发表了题为eccDNAs are apoptotic products with high innate immunostimulatory activity的研究,对上述三个基本问题做出了回答。

张毅教授实验室在表观遗传在胚胎早期发育的调控作用,做了大量杰出的工作【9-15】。早期胚胎发育过程中,基因组中大量的转座子、逆转座子会被激活。这些激活的转座子及逆转座子可能会再插入,破坏基因组稳定性。因此胚胎如何避免活性转座子、逆转座子对基因组的破坏,是一个重要的科学问题。其中一个合理的假说是:激活后转座子和逆转座子最终以eccDNA方式存在,而避免了重新插入破坏基因组。为了验证这一假说,张毅教授实验室五年前开始对eccDNA展开研究。
有效地分离和纯化研究对象,是取得研究进展的先决条件。eccDNA在生物样本中丰度极低而其环状结构又极易受损,双链上任意位点的断裂均会将环状变成线性,而导致其丢失。有效的分离纯化一直是eccDNA 研究的一大难题。传统的eccDNA分离纯化方法主要分为两步:1、碱性裂解复性粗提环状DNA;2、核酸外切酶选择性消化线性DNA。然而,文章作者发现,传统的eccDNA纯化方法远不能有效分离到满意纯度的环状DNA, eccDNA样品中60-80%的分子仍然呈线性,这与近10年来的研究报道中的结果一致。作者推测,天然生物样本中的DNA,可能存在多种修饰和复杂结构,如氧化(oxidation)、交联(cross-link),DNA复制叉、十字结(Cruciform)、脱氧鸟嘌呤四联体(G-quadruplex)等。这些复杂结构,极可能抑制了核酸外切酶的作用,使单依靠核酸外切酶消化、得到纯环状DNA,变得不可能。鉴于此,研究人员在优化传统的碱性粗提环状DNA和核酸外切酶消化线性DNA的基础上,增加了一个纯化步骤:从核酸外切酶消化产物中选择性回收环状DNA。新的方法实现了eccDNA样品在显微镜镜检时,95%以上的DNA分子呈环状(图1)。与此同时,新方法还有耗时短、稳定性高的特点:将传统耗时一周的分离纯化过程压缩到一天内完成;对平行样品、批次样品间的eccDNA纯化,表现出很好的平行性和高可再现性。
为了得到每一个eccDNA分子环的准确序列信息,研究人员还将滚环扩增(rolling cycle amplification)和第三代的纳米孔测序技术(Oxford Nanopore)结合(图1),实现了仅用三代测序获取eccDNA的全长序列。
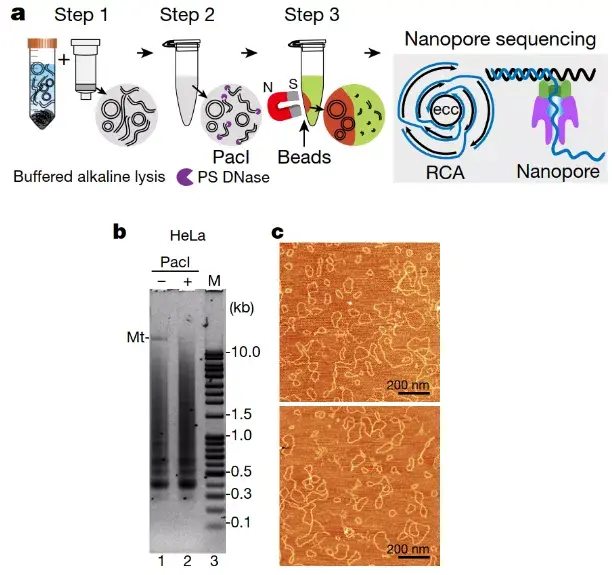
图1:(a) 新的环状DNA的纯化和测序方法;(b) eccDNA琼脂糖凝胶电泳图;(c)eccDNA分子的原子力显微镜镜检图。
为了避免普通细胞系基因组中可能存在的突变、重组、结构紊乱等因素对其eccDNA与参考基因组DNA(reference genome)序列同源比对分析的影响,作者选择从基因组相对稳定的小鼠胚胎干细胞系中提取eccDNA进行研究,获取了160多万个eccDNA分子的全长序列信息及基因组来源位置信息。分析发现,eccDNA随机来源于基因组片段,包括单片段自环化和多片段连接环化,大小集中在200bp到3kb之间,分布近似于寡聚核小体(oligo-nucleosome)间的尺寸分布规律。这些结果预示eccDNA可能是基因组DNA随机断裂产生片段的环化产物。
细胞凋亡是广泛发生于真核生物体内、和体外培养细胞中的程序性细胞死亡,其中一个重要特征就是细胞凋亡过程中,特定的核酸酶会在核小体的间区切断基因组DNA,形成寡聚核小体大小规律分布的线性片段化DNA(ladder-like)。因此作者比较了正常培养细胞和诱导凋亡的细胞中的eccDNA含量,发现凋亡细胞中的eccDNA量明显增加,说明凋亡能诱导eccDNA的发生。
为了弄清eccDNA的产生是否源于片段化的DNA,作者鉴定并敲除了小鼠干细胞中负责凋亡DNA片段化的核酸酶DNase1l3,得到了凋亡DNA片段化缺陷的细胞株(DNase1l3 KO),DNase1l3的敲除并不影响细胞凋亡导致的细胞死亡,但其凋亡寡聚核小体样的DNA片段化的现象则完全丧失,相应的是,DNase1l3 KO细胞系也失去了凋亡诱导的eccDNA发生的能力(图2);这说明eccDNA发生源于凋亡产生的DNA片段。
线性DNA片段的环化离不开DNA连接酶(DNA ligase),真核生物共有三个DNA连接酶基因:Lig1,Lig3和Lig4。通过建立DNA连接酶的单基因敲除和多基因敲除细胞系。研究人员发现,Lig1和Lig4的缺失完全不妨碍eccDNA的生成,而Lig3缺失的细胞系中的eccDNA产量则急剧降低,说明Lig3是最主要的负责DNA环化的连接酶(图2)。这回答了eccDNA是怎么发生的问题。
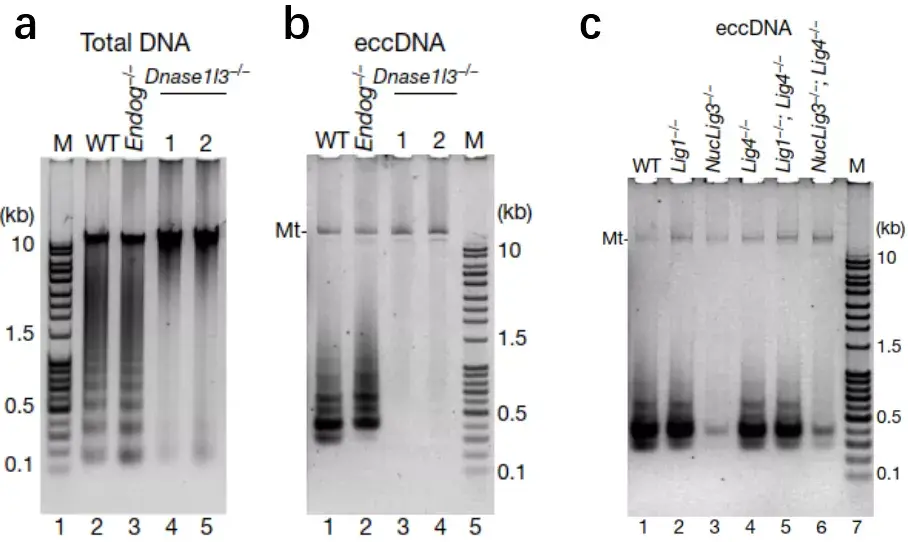
图2:(a-b)凋亡DNA片段化缺失细胞(DNase1l3 KO)不能产生eccDNA;(c)Lig3的缺失抑制了eccDNA的产生。
随后,研究人员挑战了eccDNA的第三大基本问题:eccDNA是否有功能?鉴于eccDNA在序列上没有表现出明显的特征,作者首先排除基于序列来探索eccDNA的整体生物学功能的可能。文章作者通过检阅文献发现,先天免疫(innate immunity)过程中的重要蛋白,如HMGB1,TLR9等对DNA的弯折(curvature,bending)结构表现出了更高的亲和力,因此推测,所有eccDNA分子的共性——环状结构或许在先天免疫反应中有特殊作用。
随后的研究证实了这一推测:eccDNA的环状特性赋予其超强的刺激先天免疫反应的能力。在典型的免疫细胞——树突细胞(BMDC,bone marrow derived dendritic cells) 和巨噬细胞(BMDM,bone marrow derived macrophages) 中,相比于同大小的线性基因组DNA片段, eccDNA能诱导出更高的细胞因子(图3)(Ⅰ型干扰素(IFN-α,β), 白介素6(TNF-α), 肿瘤坏死因子(TNF-α)等)的表达,显示出超强的刺激免疫反应的能力。当eccDNA分子环被单位点切断成对应的线性分子后,即失去了刺激免疫反应的超强能力(图3),证实了eccDNA的强效免疫刺激能力依赖于其环状结构。为了进一步排除eccDNA序列及潜在的转录对免疫刺激影响,作者用与eccDNA相似大小、但序列为随机生成的、人工合成环状DNA处理免疫细胞,不出预料,人工合成的DNA环完美再现了纯化的天然eccDNA对免疫细胞的超强刺激能力(图3)。深入研究显示,eccDNA的超强先天免疫刺激能力依赖于胞内Sting 信号途径而非Myd88信号途径。
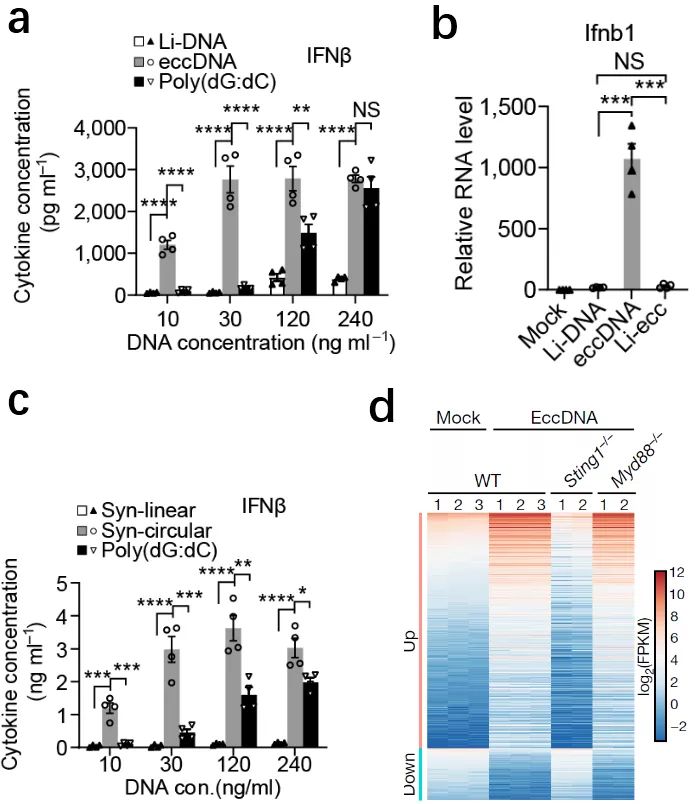
图3:(a)eccDNA具有超强的诱导干扰素IFN-β表达的能力(d)eccDNA被线性化(Li-DNA)后失去诱导干扰素IFN-β表达的能力;(c)人工合成的随机序列DNA环具有超强的诱导干扰素IFN-β表达的能力;(d)eccDNA对免疫的刺激依赖于Sting 信号途径而非Myd88信号途径。
总之,该研究就长达半个多世纪的eccDNA谜题,在三个最基本的方向上给出了清晰的答案:1,eccDNA随机来源于染色体基因组DNA,没有明显的位置或序列的特异性(从哪来),2,eccDNA是Lig3介导的凋亡DNA片段的环化产物(怎么来),3,eccDNA具有超强的刺激先天免疫反应的能力(能做什么)。该研究对细胞凋亡和先天免疫两大领域具有深远的影响,特别是对短时间内产生大量细胞死亡的多种疾病、或疾病治疗过程,如细菌、病毒的急性致命传染病、癌症的免疫治疗等。此外,该研究也对核酸疫苗和疫苗佐剂(adjuvant)的设计提供了新的设计思路和路径。
张毅教授为本文的通讯作者,王元高博士为本文的第一作者。王萌博士、Mohamed Nadhir Djekidel博士、陈欢博士、刘迪博士和Frederick Alt教授亦对本研究做出重要贡献。该研究中,原子力显微镜的使用得到了Wyss研究所的Yin Peng教授的倾情相助。
原文链接:
https://www.nature.com/articles/s41586-021-04009-w
专家点评
钟波、舒红兵(武汉大学免疫与代谢前沿科学中心)
染色体外环状DNA(eccDNA)是一类游离于染色体外的环状DNA,广泛存在于不同类型的细胞和组织中,参与调控基因的表达、细胞增殖、DNA损伤修复等过程。既往研究中分离纯化的eccDNA通常含有线性DNA的污染,干扰了对eccDNA所调控的生理过程的理解。在哈佛医学院张毅教授研究组最新发表在Nature的这项研究中,作者开发了一种获得高纯度eccDNA的方法,并发现HeLa细胞和小鼠胚胎干细胞(mES)中都含有eccDNA。mES中eccDNA大小从几百到几千个碱基(平均1kb),没有序列特异性且来源于整个基因组。进一步研究表明,mES中eccDNA的产生依赖于凋亡过程中DNA片段化的关键酶DNaseg以及DNA连接酶Lig3,表明这些eccDNA来自于基因组DNA片段的环化。诱导细胞凋亡能促进eccDNA的产生,该过程主要发生在细胞凋亡的后期。过去的研究认为细胞凋亡是一种“免疫沉默”的细胞死亡方式,凋亡过程中产生的凋亡小体将片段化的DNA包裹起来,避免使其激活天然免疫细胞的DNA受体,从而避免诱导炎症反应。张毅研究组这项最新研究表明,凋亡细胞释放的eccDNA能显著激活天然免疫细胞(如树突状细胞与巨噬细胞)表达I型干扰素以及TNF/IL-6等炎性细胞因子。这些结果表明细胞凋亡也能激活天然免疫效应。
该研究同时发现,与相同浓度和长度的线性DNA相比,eccDNA与合成的环状DNA能更为有效地激活I型干扰素与炎性细胞因子的表达,且该过程依赖于STING(我们自己实验室于2008年发现并报道了同一个蛋白并命名为MITA), 一个在DNA诱导的天然免疫信号转导中发挥关键作用的接头蛋白。线性DNA可激活DNA感受蛋白cGAS产生2’-3’cGAMP信号分子,2’-3’cGAMP结合STING/MITA并促进其活化,进而诱导下游转录因子IRF3的激活及天然免疫效应基因的表达。eccDNA是否通过cGAS激活STING/MITA尚不明确。细胞凋亡在机体发育、T/B细胞增殖分化等过程中起着极为重要的作用,eccDNA在这些过程中的潜在功能和机制有待更多研究。总之,该项研究揭示了细胞凋亡激活天然免疫的全新概念和机制,有助于我们更好的理解肿瘤、感染及炎性疾病的发生机制并研发有效的干预手段。该项成果是近年细胞生物学与免疫学领域的一项重要突破。
制版人:十一
参考文献
1 Hotta, Y. & Bassel, A. Molecular Size and Circularity of DNA in Cells of Mammals and Higher Plants. Proc Natl Acad Sci U S A 53, 356-362, doi:10.1073/pnas.53.2.356IF: 12.779 B1 (1965).2 Van Bruggen, E.F.J., Borst, P., Ruttenberg, G.J.C.M., Gruber, M. and Kroon, A.M., 1966. Circular mitochondrial DNA. , 119(2), pp.437-439.
3 Sinclair, J.H. and Stevens, B.J., 1966. Circular DNA filaments from mouse mitochondria. Proceedings of the National Academy of Sciences of the United States of America, 56(2), p.508.
4 Nass, M.M., 1966. The circularity of mitochondrial DNA. Proceedings of the National Academy of Sciences of the United States of America, 56(4), p.1215.
5 Roth, T.F. and Helinski, D.R., 1967. Evidence for circular DNA forms of a bacterial plasmid. Proceedings of the National Academy of Sciences of the United States of America, 58(2), p.650.
6 Hickson, F.T., Roth, T.F. and Helinski, D.R., 1967. Circular DNA forms of a bacterial sex factor. Proceedings of the National Academy of Sciences of the United States of America, 58(4), p.1731.
7 Shibata, Y., Kumar, P., Layer, R., Willcox, S., Gagan, J.R., Griffith, J.D. and Dutta, A., 2012. Extrachromosomal microDNAs and chromosomal microdeletions in normal tissues. Science, 336(6077), pp.82-86.
8 Turner, K.M., Deshpande, V., Beyter, D., Koga, T., Rusert, J., Lee, C., Li, B., Arden, K., Ren, B., Nathanson, D.A. and Kornblum, H.I., 2017. Extrachromosomal oncogene amplification drives tumour evolution and genetic heterogeneity. Nature, 543(7643), pp.122-125.
9 Inoue, A. and Zhang, Y., 2014. Nucleosome assembly is required for nuclear pore complex assembly in mouse zygotes. Nature structural & molecular biology, 21(7), pp.609-616.
10 Lu, F. and Zhang, Y., 2015. Cell totipotency: molecular features, induction, and maintenance. National science review, 2(2), pp.217-225.
11 Lu, F., Liu, Y., Inoue, A., Suzuki, T., Zhao, K. and Zhang, Y., 2016. Establishing chromatin regulatory landscape during mouse preimplantation development. Cell, 165(6), pp.1375-1388.
12 Inoue, A., Jiang, L., Lu, F., Suzuki, T. and Zhang, Y., 2017. Maternal H3K27me3 controls DNA methylation-independent imprinting. Nature, 547(7664), pp.419-424.
13 Matoba, S., Wang, H., Jiang, L., Lu, F., Iwabuchi, K.A., Wu, X., Inoue, K., Yang, L., Press, W., Lee, J.T. and Ogura, A., 2018. Loss of H3K27me3 imprinting in somatic cell nuclear transfer embryos disrupts post-implantation development. Cell stem cell, 23(3), pp.343-354.
14 Chen, Z. and Zhang, Y., 2020. Maternal H3K27me3-dependent autosomal and X chromosome imprinting. Nature Reviews Genetics, pp.1-17.
15 Chen, Z., Djekidel, M.N. and Zhang, Y., 2021. Distinct dynamics and functions of H2AK119ub1 and H3K27me3 in mouse preimplantation embryos. Nature Genetics, 53(4), pp.551-563.(参考文献可上下滑动阅览)